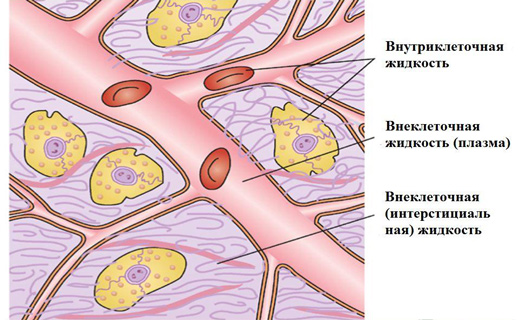
Что собой представляет объем внеклеточной жидкости в организме? Внеклеточная вода
Что собой представляет объем внеклеточной жидкости в организме?
Чтобы разобраться, какую роль играет хлорид натрия в организме, начнем с древних форм жизни - одноклеточных морских организмов. Море в данном случае выполняет следующие функции: - является питательной средой, из которой живой организм получает необходимые вещества для построения клетки и поддержания ее жизнедеятельности;
- служит неисчерпаемым резервуаром кислоты;
- играет роль клоаки, в которую выделяются отходы, образующиеся в процессе обмена веществ.
Одноклеточные организмы
Море можно расценивать и как внешнюю среду клеток вследствие постоянной концентрации в нем солей и кислоты. Одноклеточные организмы обладают способностью активно пропускать вещества через свою диафрагму. Они могут создавать такие внутренние концентрации минеральных составных частей и питательных веществ, которые значительно отклоняются от состава питательного раствора. Внутри клетки содержатся в основном ионы калия (К*), магния (Mg2*), фосфата (Р043), сульфата (БО,2), тогда как в морской воде преобладают ионы натрия (№*), кальция (Са2*) и хлора (О).
Для более высокоорганизованных живых организмов проблематичность жизни вытекает из двух фундаментальных биологических факторов: высокодифференцированной связи органов, вследствие чего все клетки практически требуют специального состава и постоянства внеклеточной жидкости, подобно одноклеточным в море; из соотношения между внутриклеточной и внеклеточной жидкостями в высокоорганизованном организме с большим преобладанием внутриклеточной жидкости. Так, например, в организме взрослого человека содержится около 30 литров внутриклеточной жидкости и лишь около 10 литров - внеклеточной. В такой ситуации может помочь лишь мощный механизм регулирования, основной задачей которого является предотвращение обеднения внеклеточного объема кислотой, питательными и минеральными веществами и избежание обогащения его продуктами разложения при обмене веществ. При этом высокоорганизованное живое существо использует особые органы, которые служат для усваивания и перемещения в организме кислоты, воды и минеральных веществ, а также выделения продуктов обмена веществ. К тому же имеется система, сравнивающая состав ионов во внеклеточной жидкости с нормальной концентрацией.
Все дело в воде
Для обеспечения жизнедеятельности клеток, чтобы могла работать система подвода питательных веществ и отвода продуктов обмена, необходим носитель - вода. Так как вследствие выделения мочи и пота организм теряет воду, требуется одновременно с приемом пищи восполнять потери воды. Причем в зависимости от внешних условий (температуры, влажности, интенсивности работы) потребность в воде для человека может резко изменяться. В Европе взрослый человек потребляет около 2 литров воды в сутки - естественно, столько же и выводится. Ниже показан баланс жидкости человеческого организма с окружающей средой.
Внеклеточная, равно как и внутриклеточная, жидкость, помимо воды, как указывалось ранее, содержит и растворенные соли, основными из которых являются хлориды натрия и калия. Следовательно; указанные жидкости - растворы и, как всякий раствор, характеризуются концентрацией соли, а поскольку вода свободно перемещается через мембрану клеток, то концентрации солей в растворах, все время выравниваются. Это происходит благодаря наличию осмотического (диффузионного) давления (Осмос от греч. osmos - толчок, давление, диффузия вещества через полупроницаемую мембрану, разделяющую растворы). После выравнивания осмотического давления растворы становятся изоосмотическими. Поэтому вне- и внутриклеточные жидкости изоосмоти.чны, хотя их ионный состав различен. Изоосмотичность и служит Тем биологическим фактором, благодаря которому осуществляется распределение воды во внутри- и внеклеточных пространствах.
Объем внеклеточной жидкости в организме
Если количество соли в организме (внеклеточной жидкости) увеличивается вследствие ее приема или уменьшения выхода, то из клетки вода станет «вытекать» до тех пор, пока не будет достигнута изоосмотичность. С увеличением во внеклеточной жидкости количества воды концентрация соли в ней понизится, и вода начнет «натекать» во внутриклеточную жидкость. Что случилось бы без этого, механизма регулирования, можно продемонстрировать в лабораторном опыте на красных кровяных тельцах. Если поместить красные кровяные тельца в гипотонический раствор, концентрация соли в котором вполовину меньше, чем во внеклеточном пространстве живого организма, они будут насыщаться водой до тех пор, пока не лопнут. Гипотонический раствор, концентрация соли в котором превышает ее концентрацию во внеклеточном пространстве, оказывает на красные кровяные тельца обратное действие: они отдают жидкость до полного сморщивания.
Вместе с тем каждый лишний грамм соли требует 120 - 130 мл воды. И, наоборот, если количество жидкости в организме сокращается, то сокращается и количество жидкости во внеклеточном отделении. Но такие сокращения, равно как и увеличения внеклеточной жидкости, могут беспоследственно проходить лишь только в определенном диапазоне, а концентрация внеклеточного натрия указывает на любое заметное отклонение от обычного ограниченного диапазона.
www.medmoon.ru
Гомеостаз жидкостей организма — SportWiki энциклопедия
Гомеостаз жидкостей организма[править]
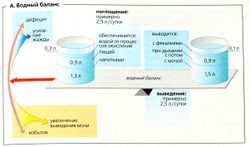
 А. Водный баланс
А. Водный баланс Жизнь не может существовать без воды. Вода является начальным и конечным продуктом бесчисленных биохимических реакций. Она служит растворителем, транспортным средством, теплоизолятором, охладителем, а также имеет множество других функций. Вода присутствует в клетках в виде внутриклеточной жидкости, а также окружает клетки в составе внеклеточной жидкости. Она обеспечивает постоянство окружающей среды (внутренней среды) для клеток организма, чем похожа на среду первичного океана, окружавшего первые одноклеточные организмы.
Объем циркулирующей жидкости в организме остается относительно постоянным, если должным образом поддерживается водный баланс (А). В среднем потребление жидкости составляет —2,5 л в сутки (в виде напитков, твердой пищи, а также продуктов метаболического окисления). Потребление жидкости должно быть достаточно высоким, чтобы компенсировать потери воды вследствие мочеиспускания, дыхания, испарения и дефекации. Средний дневной оборот воды составляет 2,5 л/70 кг (1/30 массы тела) у взрослых и 0,7 л/10 кг (1/10 массы тела) у младенцев. Таким образом, у младенцев водный баланс более подвержен отклонениям.
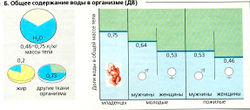 Б. Общее содержание воды в организме (ДВ)
Б. Общее содержание воды в организме (ДВ) В организме могут быть значительные увеличения оборота воды, но в норме они должны адекватно компенсироваться. Потеря воды при дыхании происходит, например, в результате гипервентиляции на большой высоте и в результате испарения при высокой температуре окружающей среды (например, пешие прогулки на солнце или особые температурные условия на работе - металлургическое производство). И то и другое может приводить к потере нескольких литров воды в течение часа, что должно быть компенсировано путем соответственного увеличения потребления жидкостей (и соли). И наоборот, увеличенное потребление жидкостей приводит к увеличению объема экскретируемой мочи.
Содержание воды в организме. Доля воды (ДВ) составляет от 0,46 (46%) до 0,75 общей массы тела (МТ = 1) в зависимости от возраста и пола (Б). У младенцев ДВ составляет 0,75, у молодых мужчин (женщин) 0,64 (0,53), а у пожилых мужчин (женщин) всего 0,53 (0,46). Различия, связанные с полом (как и индивидуальные различия), определяются в основном разным содержанием жира в теле. У молодых взрослых людей в среднем доля воды в большинстве тканей составляет около 0,73 (ср. 0,2 в жировой ткани) (Б).
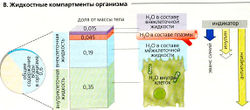 В. Жидкостные компартменты организма
В. Жидкостные компартменты организма Жидкостные компартменты. У человека в среднем ДВ —0,6, причем около 3/s ДВ (0,35 МТ) - внутриклеточная жидкость, а другие 2/s ДВ (0,25 МТ) - внеклеточная жидкость. Внутриклеточная и внеклеточная жидкости разграничены плазматической мембраной клеток. Внеклеточная жидкость находится между клетками (межклеточная жидкость, интерстиций, 0,19 МТ), в составе крови (вода в составе плазмы, 0,045 МТ) и в «трансцеллюлярных» компартментах (0,015 МТ): плевральные, перитонеальные и перикардиальное пространства, полость спинномозгового канала и мозговых желудочков, камеры глаз и просвет кишечника, протоки почек и желез (В). Плазма крови отделена от окружающих тканей эндотелием, а эпителий отделяет интерстициальное пространство от трансцеллюлярных компартментов (В). По концентрации белка плазма существенно отличается от состава остальной внеклеточной жидкости. Более того, существует фундаментальная разница в ионном составе внеклеточной и внутриклеточной жидкостей (с. 99В). Поскольку ионы Na+ в основном находятся во внеклеточном пространстве, по общему содержанию Na+ в организме можно определить объем внеклеточной жидкости (с. 176).
Измерение объемов жидкостных компартментов. В клинической медицине объемы жидкостных компартментов тела обычно измеряют с использованием индикаторов методом разбавления. При условии что индикаторное вещество S, введенное в кровоток, проникает только в компартмент-мишень (В), объем У можно рассчитать по формуле
V[л] = S [моль]/Сs [моль/л], [7.12],
где Сs - концентрация индикатора S в компартменте-мишени (определяется по анализам крови). Объем внеклеточной жидкости обычно измеряется с использованием в качестве индикатора инулина или бромида натрия (он не входит в клетки), а ДВ - с использованием антипирина, тяжелой воды (DpO) или меченой НдО. Объем внутриклеточной жидкости примерно равен разности объемов распространения антипирина и инулина. Меченый альбумин или эванс синий, который полностью связывается белками плазмы, может быть использован для измерения объема плазмы. Объем крови можно найти как отношение объема плазмы к разности [1 - гематокрит], а межклеточный (интерстициальный) объем - как разность объемов внеклеточной жидкости и плазмы. (Так как после центрифугирования оказывается, что 0,1 объема плазмы приходится на эритроциты, при расчете общего объема крови следует брать не 1, а 0,91.) Объем крови также можно измерить при введении эритроцитов, содержащих радиоактивную метку 51 Сr; тогда объем плазмы получают, умножая объем крови на (0,91 - Hct).
Регуляция содержания солей и воды[править]
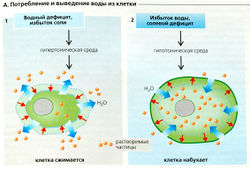 А. Потребление и выведение воды из клетки
А. Потребление и выведение воды из клетки Осморегуляция. Осмоляльность большинства жидкостей тела составляет примерно 290 мОсм/кг Н2О, так что внутри- и внеклеточная жидкости (ICF и ECF) находятся в осмотическом равновесии. Любое увеличение осмоляльности внеклеточной жидкости, например из-за абсорбции NaCI или потери воды, приводит к оттоку жидкости из клеток (сжатие клеток, А1). Падение внеклеточного осмотического давления при поглощении или введении больших объемов воды или при потере Na+ (например, при дефиците альдостерона) приводит к тому, что вода из ECF устремляется в клетку (набухание клеток, А2). И то и другое представляют риск для нормального функционирования клетки, но клетка имеет механизмы защиты.
Плазматическая мембрана клетки содержит механорецепторы, которые участвуют в регуляции баланса потоков ионов и воды - например, выход К+ и Cl- при увеличении объема клетки и вход Na+, К+ и Cl- при плазмолизе. Эти механизмы также контролируют баланс при увеличении объема клеток из-за усиленного поглощения Na+ и глюкозы в слизистой оболочке кишечника или из-за кратковременной гипоксии (со снижением активности К+-Na+-АТФазы).
Клетки, физиологическая активность которых связана со значительными перепадами осмоляльности (например, в почках), также способны регулировать собственную осмоляльность с помощью образования/абсорбции или выведения/ресорбции низкомолекулярных веществ, известных как органические осмолитики (например, бетаин, таурин, миоинозит, сорбит).
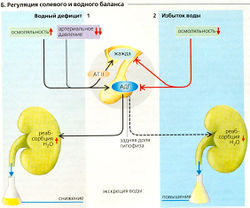 Б. Реляция солевого и водного баланса
Б. Реляция солевого и водного баланса Осмоляльность внеклеточной жидкости должна точно регулироваться, чтобы защитить клетки от больших флуктуаций объема. Осморегуляция контролируется центральными осморецепторами (или осмосенсорами), находящимися в циркумвентрику-лярных органах (SF0 и 0VLT, см. далее). Колебания объема Н2О в ЖКТ регистрируются периферическими осмосенсорами в области воротной вены и передаются в гипоталамус по афферентным нейронам блуждающего нерва.
Водный дефицит (Б1). Общая потеря воды (гиповолемия), например, из-за потоотделения, мочеиспускания или дыхания делает внеклеточную жидкость гипертоничной. Увеличение осмоляльности на 1-2% или более (= 3-6 мОсм/кг Н2О) достаточно для стимуляции секреции АДГ (антидиуретический гормон = вазопрессин) задней долей гипофиза (В1). АДГ снижает экскрецию Н2О с мочой
Однако также необходимо поступление воды в организм извне. Сходным же образом гипертоническая спинномозговая жидкость (СМЖ) через осморецепторы в сосудистом органе конечной пластинки (0VLT) и субфорникальном органе гипоталамуса (SF0) стимулирует секрецию ангиотензина II (AT II), который вызывает гиперосмотическую жажду (В). Изотоническая гиповолемия, например вследствие кровопотери или в результате гипонатриемии (Г1), также вызывает жажду (гиповолемическая жажда, В), но относительный дефицит межклеточной жидкости в этом случае больше (>10%), чем относительное увеличение осмоляльности при гиперосмотической жажде (1-2%). Рецепторы гиповолемии в основном представлены предсердными сенсорами. Через афферентные тракты и ядро одиночного тракта (NTS) запускается секреция центрального AT II в SF0 (В, И), а через периферическую нервную систему и почечные бета1-адренорецепторы активируется периферическая ренин-ATII система (А4). Падение среднего кровяного давления ниже 85 мм рт. ст. обусловливает значительное повышение уровня секреции ренина непосредственно в почке. Как и центральный, периферический AT II может вызывать жажду и повышенный Na+-аппетит, так как SF0 и 0VLT расположены за пределами ГЭБ.
Релаксин, пептидный гормон, синтезируемый желтым телом у беременных, связывается с рецепторами в SF0 и 0VLT. Он вызывает жажду и стимулирует секрецию АДГ. Несмотря на пониженную осмоляльность плазмы, которая могла бы подавлять жажду и секрецию АДГ в период беременности, релаксин обеспечивает нормальное или даже повышенное потребление жидкости в это время.
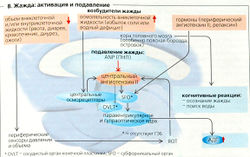 В. Жажда: активация и подавление
В. Жажда: активация и подавление Жажда - субъективное ощущение и мотивация к поиску жидкостей и питью. Жажда - реакция организма на гиперосмоляльность или гиповолемию (более 0,5% массы тела - порог жажды), что стимулирует первичное питье. Первичное питье подавляет жажду до того, как осмоляльность полностью нормализуется. Такое досрочное снижение жажды является невероятно точным механизмом, так как связано с определением объема через афферентные сигналы от волюмо- и осморецепторов горла, ЖКТ и печени. Первичное питье, однако, является скорее исключением в условиях постоянной доступности жидкости, пригодной для питья. Обычно человек пьет, потому что у него пересохло во рту или во время еды, но может и без необходимости - например, по привычке или потому, что так принято. Такое повседневное питье называется вторичным питьем.
В пожилом возрасте 30% людей 65-74 лет реже испытывают жажду, а 50% людей старше 80 лет пьют слишком мало жидкости. O возрастом способность к концентрированию мочи и секреции АДГ и альдостерона также понижается, что ведет к сопутствующему водному дефициту. Вследствие рассеянности и забывчивости потребление жидкости снижается, что порождает порочный круг. Некоторые пожилые люди стараются бороться с ночным и учащенным мочеиспусканием путем потребления меньшего количества жидкости, что также обезвоживает организм.
Избыток воды (Б2). Абсорбция гипотонической жидкости, включая, например, промывание желудка или введение раствора глюкозы (глюкоза быстро метаболизируется до СОд и НдО), снижает осмоляльность внеклеточной жидкости. Этот сигнал ингибирует секрецию АДГ, что приводит к водному диурезу и нормализации осмоляльности плазмы менее чем за 1 час.
Водная интоксикация происходит, когда избыточные количества воды абсорбируются слишком быстро, что ведет к симптомам тошноты, рвоты и шоку. Данное состояние вызывается несвоевременным падением осмоляльности плазмы до того, как происходит адекватное ингибирование секреции АДГ.
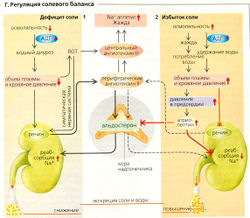 Г. Регуляция солевого баланса
Г. Регуляция солевого баланса Регуляция объема. За сутки в организме всасывается примерно 8-15 г NaCI. Почки должны одновременно экскретировать такое же количество, чтобы сохранить количество Na+ в организме на постоянном уровне и обеспечить гомеостаз внеклеточной жидкости. Поскольку Na+ является основным внеклеточным ионом (баланс Cl- поддерживается во вторую очередь), изменения содержания Na+ в организме ведут к изменению объема внеклеточной жидкости. Это регулируется в основном следующими факторами.
- Ренин-ангиотензиновая система (РАО AT II не только индуцирует жажду и солевой аппетит, но также снижает скорость клубочковой фильтрации и усиливает секрецию АДГ и альдостерона, что, в свою очередь, подавляет выведение Na+ (Г2) и, несмотря на потребление воды, устойчиво поддерживает солевой аппетит.
- Окситоцин, синтезируемый гипоталамусом нейромедиатор, ингибирует активность нейронов, которые поддерживают постоянный солевой аппетит, и посредством нервной регуляции повышает выведение NaCI.
- А триопептин (натрийуретический пептид предсердия, ANP или ПИП) - это пептидный гормон, секретируемый специальными клетками предсердия в ответ на увеличение объема внеклеточной жидкости и, следовательно, предсердного давления. ПНП подавляет жажду и понижает уровень секреции АДГ. Он способствует выведению Na+ почками путем увеличения фильтрационной фракции и ингибирует реабсорбцию Na+ в собирательной трубочке. ПНП, таким образом, функционирует как антагонист ренин-ангиотензиновой системы (РАС).
- АДГ. Секреция АДГ стимулируется путем: (а) увеличения осмоляльности плазмы и СМЖ\ (б) рефлекса Гаера-Генри, который запускается, когда волюморецепторы в предсердии предупреждают гипоталамус о снижении (>10%) объема внеклеточной жидкости (давления в предсердиях). AT II является ключевым фактором в данном процессе.
- Прессорный диурез, вызванный повышенным артериальным давлением, например, из-за увеличенного объема внеклеточной жидкости, приводит к повышенной экскреции Na+ и воды, таким образом снижая объем внеклеточной жидкости и кровяное давление. Эта система обратной связи считается основным механизмом долговременной регуляции кровяного давления.
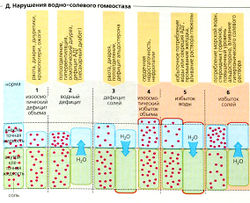 Д. Нарушения водно-солевого гомеостаза
Д. Нарушения водно-солевого гомеостаза Дефицит соли (Г1). Если гипонатриемия развивается в условиях нормального первичного содержания воды в организме, то осмоляльность крови и секреция АДГ снижаются, временно уменьшая выведение воды. Объем внеклеточной жидкости, объем плазмы и кровяное давление последовательно снижаются (И). Это, в свою очередь, активирует РАС, что вызывает гиповолемическую жажду из-за секреции AT II и индуцированного секрецией альдостерона удержания Na+ в результате. Удержание Na+ увеличивает осмоляльность плазмы, что ведет к секреции АДГ и в итоге к удержанию воды. Дополнительное потребление жидкостей в ответ на жажду также помогает нормализовать объем внеклеточной жидкости.
Избыток соли (Г2). Ненормально высокое содержание NaCI в организме, например, после потребления соленой жидкости ведет к возрастанию осмоляльности плазмы, удержанию воды организмом (жажда питье) и секреции АДГ. Таким образом, объем внеклеточной жидкости растет, и активность РАС сдерживается. Дополнительная секреция атриопептина, возможно, вместе с натрийуретическим гормоном с более долгим временем полужизни, чем у атриопептина (уабаина), ведет к увеличению выведения NaCI и Н2О и, следовательно, к нормализации объема внеклеточной жидкости.
Если осмоляльность остается нормальной, нарушения солевого и водного гомеостаза (Д1,4) воздействуют только на объем внеклеточной жидкости. Когда осмоляльность внеклеточной жидкости возрастает (гиперосмоляльность) или снижается (гипоосмоляльность), вода во внеклеточных и внутриклеточных компартментах перераспределяется (Д2, 3, 5, 6). Основные причины этих нарушений перечислены в Д (оранжевый фон). Эффектами этих нарушений являются гиповолемия в случаях 1, 2 и 3, клеточный отек (набухание) при нарушениях 3 и 5 (например, отек мозга) и интерстициальный отек (задержка воды, легочный отек) при нарушениях 4, 5 и 6.
sportwiki.to
Вода внеклеточная - Справочник химика 21
Аэробная переработка стоков —это самая обширная область контролируемого использования микроорганизмов в биотехнологии. Она включает следующие стадии 1) адсорбция субстрата на клеточной поверхности 2) расщепление адсорбированного субстрата внеклеточными ферментами 3) поглощение растворенных веществ клетками 4) рост и эндогенное дыхание 5) высвобождение экскретируемых продуктов 6) выедание первичной популяции организмов вторичными потребителями. В идеале это должно приводить к полной минерализации отходов до простых солей, газов и воды. Эффективность переработки пропорциональна количеству биомассы и времени контактирования ее с отходами. [c.249]Внутриклеточная вода Внеклеточная вода [c.64]
Поступление, распределение и выделение из организма. В организме животных и человека К. играет важную роль, участвуя в генерации биоэлектрических потенциалов, поддержании осмотического давления, участвует в углеводном обмене, синтезе белков. Он является основным внутриклеточным катионом. К. поступает в организм с пищей и водой. В организме взрослого содержится 4000—9000 мэкв К. или 160— 250 г, из них только 2 % находится во внеклеточной жидкости (интерстициальная жидкость, плазма крови). Суточная потребность в К. составляет 2—3 г у взрослых, 12—16 мг/кг у детей. Содержание К. (в мэкв) тело со скелетом 68, кости 15, зубы 17, мышцы 100, сердце 64, легкие 38, мозг 84, печень 55, почки 45, эритроциты 150, сыворотка крови 4,5 спинномозговая жидкость 2,3 лимфа 2,2. Обмен К. в организме происходит чрезвычайно интенсивно за 1 минуту в клетках мозга обменивается 3,3—4 % К- в сетчатке глаза 8—10,7%, Выведение [c.49]
В настоящее время в значительной степени изучены молекулярные механизмы реабсорбции и секреции веществ клетками почечных канальцев. Так, установлено, что при реабсорбции натрий пассивно поступает из просвета канальца внутрь клетки, движется по ней к области базальной плазматической мембраны и с помощью натриевого насоса поступает во внеклеточную жидкость. До 80% энергии АТФ в клетках канальцев почек расходуется на натриевый насос . Всасывание воды в проксимальном сегменте происходит пассивно в результате активного всасывания натрия. Вода в этом случае следует за натрием. Кстати, в дистальном сегменте всасывание воды происходит вне всякой зависимости от всасывания ионов натрия этот процесс регулируется антидиуретическим гормоном. [c.611]
По местонахождению в организме воду разделяют на две группы вода внутриклеточных жидкостей и вода внеклеточных жидкостей. [c.151]
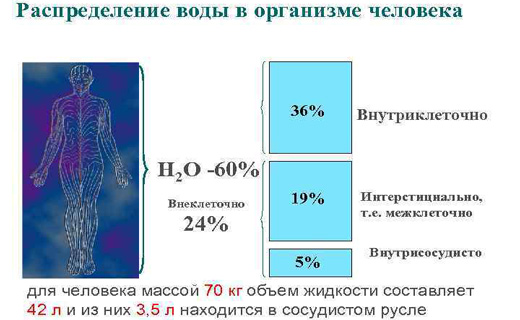
Известно, что общее содержание воды в организме человека составляет 60—65% от массы тела, т.е. приблизительно 40—45 л (если масса тела 70 кг) 7з общего количества воды приходится на внутриклеточную жидкость, 7з — на внеклеточную. Часть внеклеточной воды находится в сосудистом русле (5% от массы тела), большая часть—вне сосудистого русла—это межуточная (интерстициальная), или тканевая, жидкость (15% от массы тела). Кроме того, различают свободную воду , составляющую основу внутри- и внеклеточной жидкости, и воду, связанную с различными соединениями ( связанная вода ). [c.582]
Между животными клетками, с одной стороны, и растительными и бактериальными — с другой, имеется несколько кардинальных различий. К их числу относятся различия в среде обитания зтих клеток. Клетки животного организма погружены в специально созданную жидкую среду — кровь или лимфу. Эти жидкости в известном смысле подобны по составу древнему Океану, в котором некогда возникла жизнь (часто говорят поэтому, что животные носят в себе частицу моря). Суммарные молярные концентрации низкомолекулярных веществ во внеклеточных жидкостях животного и в цитоплазме близки. Позтому животные клетки находятся в осмотическом равновесии со средой, а их мембраны не подвергаются механическим нагрузкам за счет неравновесной диффузии воды внутрь клетки или из нее. [c.147]
Не для всех видов производственных сточных вод выделение регенераторов целесообразно. Если внеклеточная переработка очень длительна и сопоставима со скоростью потребления кислорода, то необходимость в устройстве регенераторов отпадает. [c.189]
Содержание воды в клетках достигает 65—80%. В протоплазме на каждую молекулу белка приходится около 1800 молекул воды, причем состав ее в клетках непрерывно обновляется. В зависимости от условий культивирования содержание воды в клетках может меняться. Часть воды находится в межклеточном пространстве, это внеклеточная вода, а часть воды находится в самих клетках. В свою очередь находящаяся в клетках вода может быть в свободном и в связанном с поверхностью макромолекул виде. [c.23]
Вьщеленные индивидуальные гликозаминогликаны могут содержать смесь цепей различной длины (рис. 5.5). Гликозаминогликаны как основное скрепляющее вещество связаны со структурными компонентами костей и соединительной ткани. Их функция состоит также в удержании большой массы воды и в заполнении межклеточного пространства. Иными словами, гликозаминогликаны —основной компонент внеклеточного вещества—желатинообразного вещества, заполняющего межклеточное пространство тканей. Они также содержатся в больших количествах в синовиальной жидкости-это вязкий материал, окружающий суставы, который служит смазкой и амортизатором. [c.187]
Поэтому для изучения сродства и влияния ряда флавоноидов различной структуры — агликонов и гликозидов — на мембраны клеток тканей артерий и вен крыс бьш использован метод спиновых зондов, в котором липофильный спиновый зонд 5 вводили в раствор, содержащий отрезок изучаемого сосуда. При этом зонд 5 встраивался в липидный бислой мембран клеток ткани сосудов и был недоступен для внеклеточной воды. По спектрам ЭПР определяли параметры вращатель- [c.577]
При определенных обстоятельствах микроорганизмы способны запасать органические и неорганические вещества в клетках (табл. 3.2а). Запасные вещества хранятся в полимерной форме внутри клетки. Микроорганизмы также способны превращать органический субстрат во внеклеточные полимерные вещества (ВПВ). Являются ли такие вещества запасными для микроорганизмов — это вопрос спорный, так как микроорганизмы обычно их в дальнейшем не используют. В процессе биологической очистки воды обнаружено три типа запасных веществ. [c.97]
Флокуляция с участием внеклеточных полимеров проходит по обычной схеме и заключается в образовании полимерных мостиков между микроорганизмами и бактериальными клетками активного ила. Флокуляция сопровождается увеличением количества коллоид- но-связанной воды и замедлением скорости фильтрования через пористую перегородку. - [c.110]
Согласно теории образования хлопьев активного ила, основанной на взаимодействии внеклеточных высокомолекулярных полиэлектролитов, последние наиболее интенсивно образуются в эндогенной фазе развития культуры, и в этот период после осаждения биомассы наблюдается наименьшая мутность жидкой фазы. В связи с этим в практике очистки воды и последующего сгущения суспензии активного ила важно учитывать воз- [c.68]
Количество циркулирующего альбумина зависит от общего объема плазмы. Потеря альбумина у больных с патологией почек приводит к разнице в осмотическом давлении между плазмой крови и внеклеточной жидкостью, что обусловливает отток воды из клеток во внеклеточное пространство. [c.999]
Если выпить большое количество морской воды, то общая концентрация электролитов во внеклеточной жидкости окажется гораздо выше, чем внутри клеток. Такой градиент концентрации вызывает отток воды из клеток во внеклеточное пространство. По мере обезвоживания клеток нежные внутриклеточные органеллы (например, митохондрии) съеживаются и в конце концов необратимо повреждаются. [c.999]
Соли натрия и калия. Соли натрия и калия содержатся во всех тканях, причем соли натрия главным образом во внеклеточных жидкостях—в плазме крови, лимфе, пищеварительных соках и т. д., а соли калия — в содержимом клеток. Соли натрия способствуют удержанию воды в тканях, а соли калия и кальция — удалению воды из тканей. Соли натрия влияют на рост организма. Соли калия угнетают сердечное сокращение. Нерастворимые соли кальция и магния (фосфорнокислые, углекислые и фтористые) входят в состав костей, а растворимые соли (хлористоводородные) — в состав плазмы крови и всех биологических жидкостей. Соли кальция играют боль- [c.240]
Поверхностные покрытия (краски различные типы лаков) играют двоякую роль они выполняют декоративную функцию и защищают покрываемую поверхность от вредных воздействий среды, в том числе и от микроорганизмов. Из-за постепенное отказа от введения свинца в состав красок и широкого распространения эмульсионных покрытий возникла проблема биоповреждения самих красок. Такое повреждение происходит как при хранении красок в емкостях, так и после нанесения их на поверхность и высыхания с образованием пленки. Большинство исследований в этой области направлено на создание эффективных защитных систем, которые действовали бы все то время, пока существует данное покрытие. Краски содержат пигменты, связывающие вещества, эмульгаторы, масла, смолы и смачивающие агенты они могут быть растворены в воде или в специальных растворителях. Некоторые из этих ингредиентов, например казеин, крахмал, целлюлоза и пластификаторы,, могут разрушаться микробами, а применение альтернативных, устойчивых к микробному разрушению компонентов зачастую невозможно. Развитие микроорганизмов в пленках очень сильно зависит от факторов окружающей среды температуры, влажности, наличия на поверхности питательных веществ (например, удобрений, приносимых ветром). Повреждения в емкостях часто связаны с жизнедеятельностью бактерий, но могут быть обусловлены и развитием грибов. Кроме того, в жидких эмульсионных красках могут оставаться внеклеточные ферменты, например входящие в состав целлюлазной системы эти ферменты способны снижать вязкость эмульсии. [c.241]
Высушенные до влажности около 8% дрожжевые клетки находятся в состоянии анафюза. Для сушки наиболее пригодны дрожжи плотной консистенции с содержанием внеклеточной влаги 12—17% при общей влажности 70—71%. Вода в дрожжевой клетке находится в форме адсорбционно и осмотически связанной. Адсорбционно связанная влага прочно удерживается коллоидами клетки и трудно испаряется. Потеря ее в большинстве случаев сопровождается гибелью клетки, поэтому дрожжи высушивают до влажности не. менее 8%. Осмотически связанная влага (влага набухания), так же как и внеклеточная, удаляется без нарушения структуры клетки. [c.365]
Важную группу полисахаридов составляют гликозаминогликаны, к которым относятся гиалуроновая кислота, хондроитинсульфаты и кератансульфат. Было показано, что в ориентированных пленках молекулы этих соединений в зависимости от типа присутствующих катионов могут принимать целый ряд взаимо-превращаемых конформаций [12]. Эти конформации представляют собой группу левых спиралей, упакованных антипараллельно и отличающихся в основном степенью растянутости. Наиболее сжатой является одна из конформаций гиалуроновой кислоты, в которой одна молекула закручена вокруг другой с образованием двойной спирали [13] во всех остальных случаях молекулы упакованы бок о бок . В некоторых случаях удалось детально выяснить строение молекул, что для волокнистых веществ, в отличие от кристаллических, очень трудно сделать удалось даже выявить положение молекул воды и геометрию участков молекул, координированных вокруг катионов [14]. Важными вехами на пути понимания конформационных принципов строения полисахаридных цепей стали а) первый пример установления с помощью, рентгеноструктурного анализа упорядоченной конформации разветвленного полисахарида (внеклеточного полисахарида Е. oli) это позволило предположить, что наличие ветвлений играет важную роль при ориентации боковых цепей антипараллельно основной цепи и стабилизации таким образом конформации молекул полисахарида посредством нековалентных взаимодействий [15] б) первое изучение этим же методом структуры кристаллического гликопротеина, которое показало упорядоченность конформации его углеводной части [16]. Ко времени опубликования работы [16] определение строения (F -фрагмента иммуноглобулина G) не было доведено до конца, однако уже можно было сделать ряд важных выводов, которые будут рассмотрены ниже. [c.283]
Натрий и калий встречаются в организме преимущественно в виде ионов хорошо растворимых в воде солей эти элементы содержатся во всех тканях. Характерным является наличие большого количества натриевых солей (главным образом хлоридов, фосфатов и бикарбоната натрия) во внеклеточных жидкостях — плазме крови, лимфе, пищеварительных соках, эксудатах и т. п. Соли калия, наоборот, обычно преобладают в содержимом клеток. [c.391]
Известную роль в качестве органа выделения солей играет также кожа с ее потовыми железами. Пот на 99,5% состоит из воды. В состав сухого вещества пота входят как органические соединения, например мочевина, так и неорганические соли. Основная масса неорганических солей представлена хлористым натрием — важнейшим неорганическим соединением плазмы крови и других внеклеточных жидкостей. Если принять во внимание, что человек при усиленном потоотделении может в течение суток терять несколько литров пота, то тогда станет ясно, что -420 [c.420]
В трансформации соединений фосфора, как и азотг., принимают участие организмы практически всех трофических уровней. Растворенные фосфаты (DIP) потребляются водорослями и бактериями и трансформируются в органические соединения — эфиры фосфорной кислоты. Этот органический фосфор живого вещества включается в пищевую цепь на всех уровнях. В процессе жизнедеятельности организмов выделяются фосфаты и растворенные фосфорорганические соединения (DOP), а также образуется костное взвешенное фосфорсодержащее органическое вещество — детритный фосфор (Dp). При автолизе в воду весьма быстро поступает 30—40% DOP, которые утилизируются гетеротрофными бактериями, а также гидролизуются внеклеточной фосфатазой до DIP. Кроме того, DOP, как показано в многочисленных работах, может непосредственно ассимилироваться фитопланктоном. [c.160]
Внешний гидролиз. Внеклеточные ферменты бактерий диффундируют из биопленки в толшу воды, где и происходит гидролиз. Продукты гидролиза затем диффундируют в биопленку и там разлагаются. Этот механизм был экспериментально продемонстрирован на примере крахмала, не способного диффундировать внутрь биопленки. Кинетические уравнения, описывающие этот механизм, в упрощенном виде рассматриваются ниже. [c.235]
Одна иа наиболее сложных задач на третьей стадии обработтси сточных вод путем выдерживания в лагунах - это задача выделения водорослевой клеточной ткани и большого количества внеклеточных органических веществ, вырабатываемых водорослями. [c.293]
Поступление, распределение и выведение иэ организма. В организме Н. играет важнейшую роль, являясь одним из основных элементов, участвующих в минеральном обмене, в поддержании осмотического давления, кислотно-щелочного равновесия, Б проведении нервных импульсов. Основные источники поступления Н. в организм — питьевая вода и пища. Концентрация Н. в 2100 обследованных водных системах США, снабжающих питьевой водой около половины населения страны, находится в пределах 0,4—1900 мг/л, при этом в 42 % водных систем эта величина более 20 и в 5 % — более 250 мг/л. Ежедневное количество H., поступающего в организм взрослого человека, составляет в США 1600—9600 мг (Сгаип Luft, Ganten). В организме Н. находится, в основном, во внеклеточной жидкости весь обменный Н. в организме взрослого составляет 3890 мэкв, при этом в 17,5 л внеклеточной жидкости содержится 2450, во внутренней среде, составляющей 30,3 л — 1440 мэкв. [c.40]
У морских ежей кратковременное повышение концентрации Са активирует специфические транспортные белки в плазматической мембране яйца (возможно, при участии кальмодулина), которые используют энергию, запасенную в виде трансмембранного градиента иоиов Na , для откачивания ионов Н из клетки (см. разд. 6.4.10). Отток ионов приводит к тому, что внутриклеточная величина pH возрастает с 6,6 до 7 и в дальнейшем поддерживается на этом уровне (см. рис. 14-48). Есть данные в пользу того, что именно это повышение pH индуцирует в оплодотворенных яйцах морского ежа позднюю биосинтетическую активность. Во-первых, если повысить pH в неопло-дотворенных яйцах, инкубируя их в среде, содержащей аммиак (рис. 14-52), то процессы синтеза белков и репликации ДНК заметно усиливаются даже без повышения внутриклеточной концентрации свободных ионов Са . Во-вторых, если сразу после оплодотворения поместить яйца в морскую воду, не содержащую ионов Na (так что не будет градиента Na для откачивания ионов Н ), внутриклеточный уровень pH не повышается н поздние события, связанные с активацией яйца, не наступают. Такие яйца еще можно спасти, добавив к среде аммиака тогда pH в клетке возрастает и даже прн отсутствии внеклеточного Na индуцируется синтез белков и ДНК. [c.48]
Благодаря плазмодесмам растительный организм оказывается не простой совокупностью отдельных клеток, а сложным сообществом взаимосвязанных живых протопластов. Позтому все тело растения можно рассматривать как систему, которую образуют два компартмента 1) внутриклеточный компартмент-так называемый симпласт, состоящий из объединенного множества протопластов (в том числе протопластов ситовидных трубок флоэмы) и ограниченный объединенной плазматической мембраной всех жнвых клеток, и 2) внеклеточный компартмент, или апопласг, включающий все клеточные стенки и мертвые пустые проводящие клетки ксилемы, а также находящуюся в тех и других воду (рис. 19-18). Оба компартмента имеют свои собственные транспортные системы, однако в определенных точках онн могут сообщаться между собой, а также подвергаться локальной модификации для обеспечения контроля протекающих между ними обменных процессов. [c.175]
Хотя неповреждённый барьер кровь-ткань непроницаем для больших молекул, но газы, вода, глюкоза, электролиты и аминокислоты проходят через него к нейронам и во внеклеточное пространство. Механизмы, посредством которых осуществляется этот пассаж, следующие (George M.S. et al. — 1991) [c.447]
Нарушения проницаемости барьера кровь-ткань происходят при различного рода травмах, воспалительных процессах и инсультах, что сопровождается существенными изменениями количества внутри- и внеклеточной воды, электролитов и белков. В связи с этим авторы рассматривают две группы радиоиндикаторов, пригодных для получения сцинтиграмм головного мозга. [c.447]
Измерение внутриклеточных концентраций метаболитов. Измерение концентраций промежуточных продуктов метаболизма в живой клетке сопряжено с большими экспериментальными трудностями. Поскольку клеточные ферменты катализируют быстро протекающие метаболические превращения, одна из обычных проблем при всяком экспериментальном вмешательстве в жизнь клетки связана с тем, что данные, полученные путем измерений, отражают не физиологические, а равновей1ые концентрации метаболитов. Поэтому любая экспериментальная методика будет надежной лишь в том случае, если с ее помощью удастся мгновенно подавить все ферментативные реакции в интактной ткани и тем самым предотвратить дальнейшие превращения промежуточных продуктов метаболизма. Этой цели можно достичь путем быстрого сжатия ткани между большими алюминиевыми пластинами, охлажденными жидким азотом ( —190°С) такой прием носит название фиксация замораживанием . После замораживания, мгновенно подавляющего действие ферментов, ткань растирают в порошок и ферменты инактивируют путем осаждения хлорной кислотой. Осадок удаляют центрифугированием, а прозрачную надосадочную жидкость анализируют на содержание в ней метаболитов с помощью специфических ферментативных тестов. Истинную концентрацию данного метаболита в клетке определяют расчетным путем, учитывая общее содержание воды в ткани и данные измерений объема внеклеточного пространства, В табл. 1 приведены кажущиеся внутриклеточные концентрации субстратов и продуктов реакции фосфорилирования фруктозо-6-фосфата, катализируемой фер- [c.474]
Токсическое действие морской воды. Почки человека-великолепное устройство, регулирующее удаление ионов Ыа из крови путем образования мочи, в которой содержание ионов Ка достигает 340 мМ. Однако в морской воде концентрация N3 вдвое выше той, которую способны создать почки здорового человека. Если человек пьет только морскую воду, то происходит накопление КаС1 во внеклеточной жидкости (т.е. в жидкости, окружающей клетки тела), но не во внутриклеточной жидкости. Длительное потребление морской воды приводит к смерти вследствие повреждения клеток мозга. Почему потребление морской воды на протяжении длительного времени вызывает повреждение клеток [c.778]
Полисахариды служат источником энергии и структурными компонентами клеточных стенок и внеклеточных капсул. Многие из этих полимеров, имеющие коммерческую ценность как шромышленные клеи, были получены из растительных тканей (экстракты семян и морских водорослей, древесные экссудаты щ т. п.). Способность таких полисахаридов изменять реологические свойства воды, вызывая образование геля и влияя на [c.217]
Общий объем жидкости в организме также можно определить по разбавлению инъецированного вещества подобно определению объема плазмы или внеклеточного объема. Для этого используется антипирин или Ы-аце-тил4-аминоантипирин [18—20]. Точность этого метода, однако, также снижается из-за метаболического изменения строения, связывания антипирина протеинами плазмы и аналитического предела обнаружения антипирина. Идеальным средством определения жидкости в организме является вода, меченная тритием (Н 0) [21—23]. [c.320]
Известную роль в качестве органа выделения солей играет также-кожа с ее потовыми железами. Пот на 99,5 % состоит из воды. В состав сухого вещества пота входят как органические соединения, например мочевина, так и неорганические соли. Основная масса неорганических солей представлена хлористым натрием — важнейшим неорганическим соединением плазмы крови и других внеклеточных жидкостей. Если принять во внимание, что человек при усиленном потоотделении может в течение суток терять несколько литров пота, то тогда станет ясно, что обильное потоотделение должно вызывать заметное обессоливание организма, вредно отражающееся как на общем самочувствии человека, так и на его работоспособности. Именно поэтому рабочим горячих цехов, солдатам во время продолжительных маршей и т. д. рекомендуется время от времени пить не чистую, но слегка подсоленную воду (И. П. Разенков). [c.398]
chem21.info
Состав жидкостей
Все жидкости состоят из воды и растворенных в ней веществ.
Вода.
Вода является основным компонентом человеческого организма. У взрослых мужчин вода составляет 60 % а у женщин – 55 % массы тела. К факторам влияющим на количество воды в организме относятся.
Возраст. Как правило, количество воды в организме с возрастом уменьшается. У новорожденного количество воды составляет 70 % массы тела, в возрасте 6 – 12 месяцев – 60 %, у пожилого человека 45 – 55 %. Снижение количества воды с возрастом происходит вследствие уменьшения мышечной массы.
Жировые клетки. Содержат мало воды, поэтому количество воды в организма снижается с увеличением содержания жира.
Пол. Женский организм имеет относительно меньше воды, так как содержит относительно больше жира.
Растворенные вещества
В жидкостях организма содержатся два типа растворенных веществ – неэлектролиты и электролиты.
1. Неэлектролиты. Вещества, которые не диссоциируют в растворе и измеряются по массе (например мг на 100 мл). К клинически важным неэлектролитам относятся глюкоза, мочевина, креатинин, билирубин.
2. Электролиты. Вещества которые диссоциируют в растворе на катионы и анионы и их содержание измеряется в миллиэквивалент на литр [мэкв/л]. Электролитный состав жидкостей представлен в таблице.
Таблица 29.1
Основные электролиты жидкостных компартментов организма (приведены средние значения)
| Содержание электролитов, мэкв/л | Внеклеточная жидкость | Внутриклеточная жидкость | |
| плазма | интерстициальная | ||
| Na+ K+ Ca2+ Cl- PO43- HCO3- | 140 4 5 105 2 27 | 140 4 2,5 115 2 30 | 10 150 0 2 35 10 |
Основными внеклеточными катионами являются Na+, Са2+, а внутриклеточными К+, Мg2+. Вне клетки преобладают анионы СL-, НСО3-, а главным анионом клетки является РО43-. Внутрисосудистая и интерстициальная жидкости имеют одинаковый состав, так как эндотелий капиляров свободно проницаем для ионов и воды. Различие состава внеклеточной и внутриклеточной жидкостей обусловлено:
Непроницаемостью клеточной мембраны для ионов;
Функционированием транспортных систем и ионных каналов.
Характеристики жидкостей
Кроме состава, важное значение имеют общие характеристики (параметры) жидкостей. К ним относятся: объем, осмоляльность и рН.
Объем жидкостей.
Объем жидкости зависит от количества воды которая присутствует в данный момент в конкретном пространстве. Однако вода переходит пасивно, в основном за счет Na+.
Жидкости взрослого организма имеют объем:
Внутриклеточная жидкость – 27 л
Внеклеточная жидкость – 15 л
В том числе:
Интерстициальная жидкость – 11 л
Плазма – 3 л
Трансцеллюлярная жидкость – 1 л.
Вода, биологическая роль, обмен воды
Вода в организме находится в трех состояниях:
Конституционная (прочно связанная) воды, входит в структуру белков, жиров, углеводов.
Слабосвязанная воды диффузионных слоев и внешних гидратных оболочек биомолекул.
Свободная, мобильная вода, является средой в которой растворяются электролиты и ниэлектролиты.
Между связанной и свободной водой существует состояние динамического равновесия. Так синтез 1 г гликогена или белка требует 3 г Н2О которая переходит из свободного состояния в связанное.
Вода в организме выполняет следующие биологические функции:
Растворитель биологических молекул.
Метаболическая – участие в биохимических реакциях (гидролиз, гидратация, дегидратация и др.).
Структурная – обеспечение структурной прослойки между полярными группами в биологических мембранах.
Механическая – способствует сохранению внутриклеточного давления, формы клеток (тургор).
Регулятор теплового баланса (сохранение, распределение, отдача тепла).
Транспортная – обеспечение переноса растворенных веществ.
studfiles.net
Причины, приводящие к нарушению распределения воды и электролитов между внеклеточным и клеточным секторами.
Перемещение жидкости между клеткой и интерстицием происходит в основном по законам осмоса, т.е. вода идет всегда в сторону более высокой осмотической концентрации. Избыточное перемещение жидкости в клетку происходит в двух случаях:
1) когда во внеклеточном пространстве низкая осмотическая концентрация. Это может быть при избытке воды и дефиците соли
2) когда повышается осмос в самой клетке. Это может быть, прежде всего, при нарушении Na/К насоса. И тогда Na, движущийся в клетку по законам диффузии, хуже удаляется из клетки. Функция Na/К насоса нарушается при снижении энергообеспеченности клеток, прежде всего, из-за гипоксии и нехватки энергетических субстрактов, при отравлениях. Осмотическая концентрация в клетках может нарастать при сахарном диабете за счет образования сорбитола (сорбитоловый путь окисления глюкозы). Поэтому при регидратации больных сахарным диабетом у них легко развивается набухание клеток мозга.
Избыточное перемещение воды из клетки происходит лишь тогда, когда в интерстиции нарастает осмос (это или в силу дефицита воды, или при избытке Na, мочевины, глюкозы, этил. спирта и др. частиц).
Причины, приводящие к нарушению распределения жидкости между внутрисосудистым пространством и интерстицием.
Стенка капилляра свободно пропускает воду, электролиты и некрупномолекулярные вещества и почти не пропускает белки. Поэтому концентрация электролитов по обе стороны сосудистой стенки практически одинакова и не играет роли в перемещении воды. А вот белка значительно больше в сосудах. Они определяют онкотическое давление плазмы, которое тоже является осмотическим и поэтому, по законам осмоса, удерживает воду в сосуде и определяет движение жидкости из интерстиция в сосуд в том случае, когда гидростатическое давление крови будет меньше онкотического. Такая ситуация в норме складывается в венозной части капилляра, поэтому здесь жидкость поступает в сосуд из интерстиция.
В артериальном конце капилляра в норме гидростатическое давление превышает онкотическое, что определяет переход жидкости из сосуда в интерстиций. Механизм обмена жидкости между сосудистым руслом и интерстицием называется механизмом Старлинга. О нем подробно прочитаете на стр. 226— по новому уч. Адо (1994г).
Изменение ведущих сил (гидравлического и онкотического давления в сосудах) может нарушить процесс обмена жидкости между этими секторами. Увеличение гидравлического давления (при сердечной недостаточности) в венозном конце капилляра затрудняет возврат жидкости. Снижение его при кровопотере усиливает возврат. Это является одним из механизмов восстановления объема крови при кровопотере. Увеличение онкотического давления, например, при обезвоживании, ведет, прежде всего, к усилению возврата (это один из механизмов компенсации, восстанавливающий объем жидкости в кровеносном русле при обезвоживании). Снижение онкотического давления (при циррозе печени) ведет к усилению выдоха и затруднению возврата.
КЛАССИФИКАЦИЯ НАРУШЕНИЙ ВОДНО-СОЛЕВОГО ОБМЕНА.
Дисгидрии: дегидратации, гипергидратации и ассоциированные.
Дегидратации: вне-, внутриклеточные, общие; а также гипертонические, изотонические, гипотонические.
Гипергидратации: вне-, внутриклеточные, общие; а также гипертонические, изотонические, гипотонические.
Ассоциированные: внеклеточная дегидратация с внутриклеточной гипергидратацией; внеклеточная гипергидратация с внутриклеточной дегидратацией.
Гипергидратации
Внеклеточная гипергидратация.
Изолированно она может возникать только в том случае, если осмотическое давление во внеклеточном пространстве не изменится, т.е. задержка воды и солей будет в эквивалентных количествах. Поэтому ее еще называют изотонической гипергидратацией. Основным клиническим проявлением внеклеточной гипергидратации являются отеки изолированные или генерализованные (распространенные).
Отек — это патологический процесс, основой которого является избыточное скопление внеклеточной жидкости в интерстициальном секторе в ограниченном участке тела или генерализованно (т.е. во всем организме). Скопление жидкости в серозной полости называют водянкой.
ПАТОФИЗИОЛОГИЯВОДНО-СОЛЕВОГО ОБМЕНА.
Лекция № 2.
Патогенетические факторы отеков.
1. Повышение гидростатического давления в венозном капилляре (сердечная недостаточность, тромбофлебиты и т.д.). При этом затрудняется возврат жидкости в сосуд.
2. Снижение онкотического давления плазмы крови (заболевания печени, почек, голодание). В этом случае усиливается выход жидкости в артериальном конце капилляра.
3. Повышение проницаемости капиллярной стенки. Это облегчает выход белка из сосудов в интерстиций, где повышается онкотическое давление, что ведет к затруднению резорбции жидкости из интерстициального пространства.
4. Повышение осмотического давления в интерстиции (при воспалении).
5. Недостаточность лимфооттока —как абсолютная (сдавливание лимфатических сосудов, их закупорка, спазм), так и относительная —когда процессы транссудации превышают возможность лимфатических сосудов отводить жидкость из интерстиция. Лимфатические сосуды удаляют не только жидкость, но и белки из интерстиция, попавшие туда в избытке через поврежденную сосудистую стенку. В межклеточном пространстве повышается онкотическое давление, что способствует накоплению интерстициальной жидкости.
Развитие распространенных отеков возможно только при выраженном положительном водно-солевом балансе, т.е. при условии избытка воды и солей, за счет усиленной их задержки с участием вазопрессина и альдостерона.
6. Патогенетический фактор — неадекватная гормональная регуляция водно-солевого обмена, ведущая к накоплению изотонической жидкости.
studfiles.net
Содержание электролитов

Плазма = 3 л Интерсти- (7% от 00В) циальная жидкость
=8,5 л (20%
от ООВ) ФОВЖ — функциональный объем внеклеточной жидкости.
Внеклеточная жидкость. Из катионов внеклеточной жидкости преобладает натрий. Общее содержание его в организме составляет около 4500 мэкв (103 г) *, из которых около 2800 мэкв участвуют в обмене электролитов (остальное количество включено в кристаллические компоненты костной ткани) [3]. Концентрация натрия во внеклеточной жидкости составляет в норме 135— 145 мэкв/л(135—145ммоль/л). Кроме натрия во внеклеточной жидкости в значительных количествах присутствуют: калий 3,5— 5 мэкв/л(3,5—5ммоль/л), магний1,5—2,5мэкв/л, (0,8— 1,3 ммоль/л), кальций 4,5— 5,5 мэкв/л(2,3—2,7ммоль/л). Для сравнения — концентрация водородного иона во внеклеточной жидкости составляет только3,5—4,5Х10-5 мэкв/л(3,5—4,5ХХ10-5 ммоль/л), однако поддерживать ее на этом уровне необходимо с той же точностью, что и перечисленных выше катионов. Основными анионами внеклеточной жидкости являются ион хлорида95—106мэкв/л(95—106ммоль/л) и ион бикарбоната 22— 28 мэкв/л(22—28ммоль/л).
Внутриклеточная жидкость. Основными катионами внутриклеточной жидкости являются калий и магний, их концентрации составляют соответственно 160 мэкв/л (160 ммоль/л) и 25 мэкв/л (12,5 ммоль/л), концентрация натрия составляет лишь около 10 мэкв/л (10 ммоль/л). Основными анионами внутри клетки являются фосфат (около 100 мэкв/л) и сульфат (около 20 мэкв/л). Содержание бикарбоната и хлорида вместе равняется примерно 10 мэкв/л. Суммарный отрицательный заряд внутриклеточных белков составляет 55 мэкв/л, именно их присутствие определяет большую часть различий в ионном составе вне- и внутриклеточной сред. Как известно,10—15%внутриклеточных ионов осмотически неактивны, они предположительно связаны с внутриклеточными липттдами, белками или нуклеиновыми кислотами.
Осмотическое давление, осмолялъность и концентрация солей
Для изучения водно-электролитногогомеостаза в организме крайне важно понятие осмотического давления, физической сутью которого является стремление молекул воды проник нуть через полупроницаемую мембрану из более разведенного раствора в более концентрированный. Количественно осмотическое давление оценивается по градиенту гидростатического давления, способному предотвратить перемещение воды. Осмотическая концентрация может быть выражена в единицах осмоляльности или осмолярности. Одноосмоляльный раствор содержит 1 осмоль растворенного вещества в
1 кг воды; одноосмолярный раствор содержит 1 осмоль вещества в 1 л водного раствора. Общее количество осмолей вещества в растворе равно сумме числа молей неиони-зированноговещества и числа эквивалентов каждого иона, разделенных на их валентность. В клинике чаще пользуются понятием «осмолярности», хотя термин «осмоляльность», вероятно, более точен, так как лабораторные измерения, как правило, выражены в миллиосмолях на 1 кг. В практике разница между этими двумя понятиями невелика.
* Формула перевода миллиграммов в миллиэквиваленты:
мг
мэкв== ——————X валентность. мол. масса
Внорме осмоляльность внеклеточной и внутриклеточной жидкостей составляет приблизительно 285—295мосм/кг. Соли натрия обеспечивают90—95%осмоляльности плазмы иинтерстициаль-нойжидкости, а внутри клетки осмотическое давление обеспечивается в основном солями калия. Осмотическая концентрация внутриклеточной жидкости постоянно подвергается очень точной гомеостатической регуляции.
Все растворы одинаковой осмоляльности являются изоосмотическими. Изотоническим называют раствор, который физиологически изоосмотичен с внутриклеточной жидкостью; если внеклеточную жидкость заменить этим раствором, то перемещения воды ни внутрь клетки, ни вне ее не происходит. Например, 1,8% раствор мочевины изоосмотичен, но не изотоничсн, так как диффундирует сквозь клеточную мембрану. Изотонический раствор должен быть педиффундируемым, как 0,9% раствор хлорида натрия.
Вуравновешенном состоянии не может существовать существенного различия осмоляльности по обе стороны водопроницаемой клеточной мембраны. Если внеклеточная жидкость становится гипоили гипертонической, то происходит перемещение воды в клетку или из клетки до тех нор, пока осмотические концентрации не уравновешиваются. Однако лишь молекулы воды могут свободно проникать сквозь клеточную мембрану для восстановления осмотического равновесия, ионы натрия и калия этой возможности не имеют. Поэтому при снижении осмоляльности плазмы объем внутриклеточной жидкости увеличивается за счет уменьшения функционального объема внеклеточной жидкости независимо от величины последнего. Если осмоляльпость плазмы высокая, то объем внутриклеточной жидкости уменьшается независимо от объема внеклеточной жидкости. Термины «дефицит жидкости» или «перегрузка жидкостью» в их клиническом применении относятся только к объему внеклеточной жидкости, объем внутриклеточной жидкости в то же время может изменяться совершенно в противоположном направлении. Между осмотическим давлением внутрисосудистой и интерстициальной жидкости имеется небольшой градиент, связанный с более высоким содержанием белков во внутрисосудистом пространстве. Этот градиент называют коллоидно-осмотическимдавлением, или онкотическим давлением, оно со-
ставляет около 28 мм рт. ст. Этот градиент препятствует чрезмерной потере жидкости сквозь стенки капиллярной сети. В условиях дефицита белков онкотическое давление плазмы снижается.
Механизмы гомеостаза в норме
Почечные механизмы регуляции водно-электролитногобаланса
Поддержание постоянного объема и состава внутренней жидкостной среды организма является одной из основных функций почек. Поэтому почки, составляющие только 0,4% от массы тела, получают 20—25%минутного объема сердца или приблизительно 1200 мл/мин крови. За 24 ч 2 млн. почечных клубочков фильтруют около 160 л жидкости, содержащей 24000 мэкв натрия, 700 мэкв калия, 5000 мэкв бикарбоната и 20000 мэкв хлорида. За время, пока гломерулярный фильтрат протекает черезпрокси-мальныеотделы почечных канальцев, его объем уменьшается приблизительно на 80% за счет активной реабсорбции натрия и пассивной реабсорбции хлоридов и воды (рис. 59). Поскольку вода свободно диффундирует через эпителий проксимальных канальцев, жидкость в просвете канальцев сохраняет изоосмотичность с плазмой. Около 90% выделенного клубочками бикарбоната всасывается в проксимальных канальцах, предварительно соединившись с активно секретируемым водородным ионом и образовав угольную кислоту; последняя быстро дегидратируется до двуокиси углерода за счет действия карбоангидразы, содержащейся в ресничках эпителия проксимальных капальцев. Практически весь фильтрованный клубочками калий реабсорбируется в проксимальных канальцах, то же происходит с глюкозой и большинством амипокислот. Фракция воды и электролитов, реабсорбируемая в проксимальных канальцах, остается относительно постоянной, несмотря на изменения скорости клубочковой фильтрации. Это явление, называемое «клубочковоканальцевым равновесием», предотвращает значительные колебания натрия в организме при изменении скорости клубочковой фильтрации.
При прохождении оставшейся части фильтрата по петле Генле имеют место два важных процесса, отражающие концептрационно-фильтрационнуюфункцию почек:во-первых,в восходящем колене петли Генле канальцевая жидкость становится гипотонической по отношению к плазме за счет активного транспорта натрия (и, возможно, хлорида) через канальцевый эпителий, который на данном участке обладает уникальным свойством водонепроницаемости;во-вторых,при прохождении капальца через мозговой и сосочковый слой почечной паренхимы устанавливается осмотический градиент между канальцевым фильтратом и интерстицием за счет анатомического строения петли Генле и сопровождающих ее капилляров. Осмоляльность интерстиции папиллярного слоя почечной паренхимы у здорового молодого человека может достигать1200—1300мосм/кг, т.е. в4—5раз превышать осмоляльность плазмы. Механизм, за счет которого устанавливается и поддержи-

вается этот градиент, обычно называют «противоточным умножением» и «противоточным обменом».
Рис. 59. Схема пассивного и активного перемещения воды и ионов в нефроне в процессе образования гипертонической мочи. Концентрации канальцевой мочи и интерстициалъной жидкости даны в мосм/л, крупные цифры в рамках указывают, сколько процентов клубочкового фильтрата осталось в канальце на каждом уровне. (Pitts R. F.: Physiology of the kidney and Body Fluids, ed. 3. Chicago, 1974).
В дистальном отделе извитых капальцев содержимое уже является гипотоническим по отношению к окружающей интерстиции коркового слоя, а его объем составляет около 15% от начального объема клубочкового фильтрата. Состав этой жидкости уже в значительной степени независим от объемов и осмотических свойств жидкостной среды данного больного, в последующем канальцевый фильтрат может изменять свой состав и объем по-раз-ному, в зависимости от активности многочисленных механизмов регуляции. Ионы натрия могут реабсорбироваться или экскретироваться преимущественно в обмен на ионы калия. Ионы водорода при необходимости секретируются эпителием, в основном в виде титруемых кислот или аммония. Вода почти целиком может пассивно реабсорбироваться в гипертонической интерстиции мозгового слоя из собирательных канальцев, если эпителий последних становится проницаемым под действием антидиуретического гормона (АДГ). И наоборот, в отсутствие АДГ эпителий собирательных канальцев непроницаем для воды, поэтому почти вся вода, достигшая дистальных канальцев, выделяется. В результате описанных процессов обеспечивается поддержание объема, осмоляльности и состава жидкостей организма в весьма тес-
ных границах нормы, несмотря на довольно вариабельные диетические и метаболические условия.
Поддержание функционального объема внеклеточной жидкости (натриевая регуляция)
Функциональный объем внеклеточной жидкости является одним из наиболее тщательно поддерживаемых параметров водно-электролитногобаланса. Стабильность его обеспечивается в первую очередь регуляцией выделения натрия. Уменьшение внутрисосудистого объема путем активации барорецепторов артериальной стенки (и, возможно, левого предсердия) приводит к задержке натрия. При этом включаются другие механизмы, призванные поддерживать нормальную осмоляльность. В любой ситуации в организме действует несколько процессов, регулирующих экскрецию натрия; их иерархия еще полностью не выяснена. Вот краткое описание некоторых из этих механизмов.
Выделение альдостерона. В присутствии альдостерона натрий на уровне дистальных канальцев замещается калием. Первым этапом этого сложного процесса является секреция ренина юкстагломерулярным аппаратом почки, вероятно, в результате сниженной перфузии почки или уменьшенного поступления натрия к зоне плотного пятна в дистальных канальцах. Ренин является медиатором превращения циркулирующего в крови ангиотензиногена вангиотензин-1,который, проходя через легкие, превращается вангиотензин-II— мощный стимулятор секреции альдостерона. Кроме того, полагают, что секреция альдостерона в ответ на снижение внутрисосудистого объема и/или объема внеклеточной жидкости может непосредственно стимулироваться барорецепторами каротидного синуса, а возможно, и барорецепторами других зон.
До недавнего времени считали, что альдостерон является основным фактором регуляции натриевого баланса. Однако замечено, что в случаях, когда уровень циркулирующего в организме альдостерона остается постоянным и не подвержен, естественно, механизму обратной связи, регуляция экскреции натрия тем не менее сохраняется на хорошем уровне. Таким образом, другие факторы наряду с секрецией альдостерона могут играть столь же важную роль в регуляции выделения натрия.
Изменение гломерулярной фильтрации. Предполагалось, что изме-
нение скорости клубочковой фильтрации может вызывать параллельное изменение экскреции натрия. Действительно, значительные колебания скорости клубочковой фильтрации могут влиять на экскрецию натрия, но маловероятно, что этот механизм достаточно точен для того, чтобы претендовать на роль основного фактора регуляции натриевого баланса.
Перераспределение внутрипочечного кровотока. Эксперименты продемонстрировали, что перераспределение кровотока между кортикальными и юкстамедуллярными нефронами вызывает изменение экскреции натрия [7]. У юкстамедуллярных нефронов более длинные проксималь-
ные канальцы и петля Генле по сравнению с корковыми нефронами, поэтому они лучше приспособлены к задержанию натрия. Создается впечатление, что при натрийдефицитных состояниях перфузия почки переключается преимущественно на этот тип нефронов. В состояниях тяжелой гиповолемии кровоток коркового слоя почки может снижаться до критического уровня [8]. К сожалению, пока недостаточно данных о том, какие нервные, гуморальные, внутрипочечные механизмы или их сочетания регулируют перераспределение кровотока внутри почки [9].
Гидростатическое и онкотическое давления вокруг канальца. Фи-
зические изменения околоканальцевого пространства могут существенно влиять на экскрецию натрия [61. В частности, снижение внутрикапиллярного гидростатического давления или повышение коллоидно-осмотическогодавления в капилляре способствуют реабсорбции натрия и воды через канальцевый эпителий. Это представление подтверждается клиническими наблюдениями, в которых при коррекции гиповолемии кристаллоидными растворами отмечается больший диурез, чем при переливании плазмозаменителей [10].
Натрийуретический гормон. В течение нескольких последних лет исследователи предполагают существование гуморального фактора «неальдостероновой» природы, регулирующего экскрецию натрия. Эти предположения отчасти подтверждаются данными экспериментальных исследований [11]. Считают, что увеличение внутрисосудистого объема посредством одного или нескольких барорецепторов может стимулировать выделение натрийуретического гормона, который ингибирует реабсорбцию натрия в проксимальных канальцах. Однако данный фактор еще не выделен и его существование остается предметом дискуссий.
Поддержание осмолялъности (водная регуляция)
В противоположность недостаточно изученным и несомненно сложным механизмам регуляции функционального объема внеклеточной жидкости управление осмоляльностью жидкостей в организме осуществляется одним веществом, антидиуретическим гормоном (АДГ) [1, 12]. Если осмоляльность повышается более чем на 2%, задняя доля гипофиза начинает секретироватъ АДГ. Последний воздействует на стенку собирательных канальцев и в меньшей степени на стенку дистальных канальцев, делая ее проницаемой для воды, которая таким образом проникает в гипертоническую среду интерстиции мозгового слоя. По прямым сосудам эта жидкость возвращается в венозные сосуды почки, в результате выделяется концентрированная моча в малом количестве. Секреция АДГ продолжается до тех пор, пока осмоляльность плазмы за счет задержки воды не снизится до нормы. При уменьшении осмоляльности, наоборот, секреция АДГ угнетается. Дистальные канальцы и собирательные протоки становятся непроницаемыми для воды, реабсорбция ее прекращается или становится незначительной, в результате свободная жидкость выделяется с мочой, а осмоляльпость плазмы увеличивается. Предполагают, что повышение осмоляльности «включает» секрецию АДГ посредст-
вом уменьшения клеточного объема нейронов супраоптических ядер гипоталамуса.
«Неосмотическая» регуляция секреции АДГ. Известно, что многие неосмотические факторы стимулируют или угнетают выделение АДГ, некоторые из них играют важную роль в поддержании гомеостаза у хирургических больных [12, 13]. Наиболее существенный из этих механизмов — выделение АДГ в ответ на уменьшение объема внеклеточной жидкости и/или плазмы без изменения их осмоляльности. Секреция АДГ в условиях гиповолемии сохраняется, даже несмотря на значительное снижение осмоляльности плазмы. Хотя благодаря такой задержке свободной воды функциональный объем внеклеточной жидкости может быть восстановлен до нормы, сопутствующая этому гипопатриемия может иметь потенциально опасные последствия. Считают, что «неосмотическая» секреция АДГ стимулируется посредством барорецепторов каротидного синуса (система высокого давления) или объемных рецепторов левого предсердия (система низкого давления) .
Многие препараты, используемые при анестезии включая наркотики, барбитураты, ингляциопные апестетики, оказывают АДГ-подобноедействие [12, 13]. Неизвестно, действуют эти препараты непосредственно на гипофи-зарно-адрепаловуюсистему или же антидиуретический эффект обусловлен их гемодинамическим действием; последнее более вероятно. Боль, эмоциональное напряжение, дыхание с положительным давлением на вдохе, бета-адрено-миметическиесредства типа изопротеренола, холиномиметические средства типа ацетилхолина также стимулируют секрецию АДГ. В послеоперационном периоде нельзя недооценивать роль неосмотическнх факторов в повышенной секреции АДГ.
Регуляция содержания калия
За сутки почечные клубочки фильтруют 700 мэкв калия, который почти полностью активно реабсорбируется в проксимальпых канальцах. В дистальных канальцах калий, вероятно, пассивно секретируется в просвет канальца в обмен на натрий в соответствии с трансэпителиальньтм электрическим градиентом. После приема калия в значительных количествах выделение альдостерона обеспечивает повышенную канальцевую секрецию калия. Канальцевая секреция калия усиливается также в условиях алкалоза, причиной этого является, вероятно, изменение электрических градиентов, а не конкуренция ионов калия и водорода в отношении единственного общего экскреторного канала [1]. Кроме того, повышение канальцевой секреции калия встречается в случаях, когда в дистальных канальцах присутствует натрий в больших количествах (например, если на фоне приема диуретиков блокируется реабсорбция натрия в проксимальных канальцах и петле Генле). Обычно за сутки через почки выделяется около 20 мэкв калия, это относится также и к калийдефицитным состояниям. У страдающих заболеваниями почек обмен калия остается нормальным [14] до тех пор, пока функция почек серьезно не нарушена.
studfiles.net

















